Marine
Protected Areas for Sustainable Fisheries Management: Layang Layang Reef as a
Source of Larvae in the South China Sea
Steve Oakley and Nicolas
Pilcher[1]
Abstract
Overfishing, particularly recruitment overfishing, and ecosystem
destruction are common and serious problems in the South China Sea. Direct
intervention methods to control fishing such as controlling type and size of
gear and having closed-seasons require high levels of enforcement and have not
proven to be very successful. In
contrast, establishing marine reserves, an indirect intervention, have been
shown to be a viable tool in the fishery management armoury. Marine reserves protect spawning stocks,
maintain a natural population age structure, provide a source of recruitment,
as well as encourage adult emigration from protected to fished areas. In addition to these effects, they maintain
biodiversity, protect habitats, and allow natural ecosystem processes to
occur. In terms of management, marine
reserves are advantageous because they are simpler to enforce and they act as
an insurance against failure of other management techniques. This paper also reports on the results on
the export of larvae from Layang Layang, a protected coral reef in the Spratlys
in the South China Sea. Small reefs like
Layang Layang are valuable as a source of larvae for other reef in the
region. Indeed, marine parks are
important for sustainable fisheries and it is recommended that more parks will
be established.
Introduction
Fisheries on coral
reefs are multispecific and multigear.
It can be very productive, but typically, there is little or no
effective management (e.g., Carpenter and Alcala, 1977; Alcala, 1991;
Cabanban and Biusing, this volume). As
a result, around the South China Sea, there is widespread overfishing,
destruction of ecosystem, and declining yields (e. g., Chou et al..,
1994; McManus, 1997; Chou, 1997). In
the coastal waters of the Philippines area generally overfished to the point of
having more than twice as much as fishing pressure as necessary for optimal
harvests (McManus, 1995). Many reefs
are badly damaged from dynamiting or fish bombing and characterised by low
species richness and low biomass of commercially important species (Rubec,
1988). Reefs are dying as a result of
fishing with cyanide (Johannes and Riepen, 1995). In Indonesia, destructive fishing (Pet-Soede and Erdmann, 1998)
is damaging coral reefs. These effects
on the reef are typical of many reefs in the region (McManus, 1997). Thus, proper management is acutely needed.
A number of
management techniques have been tried but limited resources, large areas,
dispersed landing sites, and poor fishers contribute to the difficulty in
enforcement. Conventional methods, when
applied to coral reef fisheries management is inappropriate because they
require vast information on the biology of stocks and a huge budget for
implementation and are difficult to enforce (Alcala and Vande Vusse, 1994;
Berkes, 1994; Roberts and Polunin, 1993).
A non-conventional method is the establishment of marine reserves - a
method that is considered a viable alternative world-wide (Dixon, 1993; Dugan
and Davis, 1993). Marine reserves offer protective management which has several
benefits (Alcala, 1988), including the protection of spawning stocks,
protection of biomass that provide larvae to replenish distant fishing grounds,
and protection of juveniles and adults that emigrate to adjacent, unprotected
areas. In contrast, marine reserves
require minimal information on the biology of stocks and enforcement is
relatively easy. However, the
effectiveness of this approach depends on the agreement of a functional
majority of the resource-users (Alcala and Vande Vusse, 1994; Berkes, 1994).
Despite these
benefits from marine parks that were established around the world, very few
studies have quantified these benefits for fisheries management (see review
below). The Coastal Zone Management
Programme at Universiti Malaysia Sarawak (UNIMAS) is researching into the
benefits of reserves so that these advantages can be used to convince
resource-users that there are real benefits to non-destructive fishing and to
protection of areas. In particular, it
has conducted a preliminary investigation on the benefit of a protected area,
the Layang Layang in the Spratlys, in being a source of larvae that are
exported by currents to other reefal areas.
The results of this study demonstrate the value of small reefs as
sources of larvae for other reefs in the South China Sea.
Importance and Benefits of Marine Parks
Marine reserves are
important to fisheries management. They
maintain biomass of large marine organisms and genetic biodiversity. They provide biomass of individuals to
migrate to non-protected areas where they get recruited into the fishery. These benefits are reviewed and discussed
below with reference to fisheries and aquaculture.
Protection of Large Marine Organisms and Genetic biodiversity
Large fishes that
live on coral reefs such as groupers, snappers, emperors, and wrasses have
either disappeared or become very rare due to overfishing in the South China
Sea and in many coral reef areas in the Caribbean. These large, solitary, and frequently territorial fishes are at
risk from simple artisanal overfishing and intensive capture of fishes for the
live-fish trade in Southeast Asia (see related articles in this volume). The live fish trade has swept through most
reefs in the South China Sea in an effort to satisfy the Hong Kong driven demand
for large rare fish. Incentives are
high when wholesale prices (in 1995) vary from $ 30-40 per kg for ordinary
groupers, $ 40-60 kg for red-coloured groupers, and $60-90 for Napoleon wrasse
(Cheilinus undulatus) or Polka dot Grouper (Cromileptes altivelis). Due to the live-fish trade, these fishes are
currently classified as vulnerable although in many areas they are locally
extinct although they have very wide ranges.
This has prompted the inclusion in early 1996 of some of the largest
fish species, Napoleon wrasse and bumphead parrotfish into the IUCN Red List.
The biggest of these fishes can only be conserved and protected
effectively from overfishing by the use of protected areas. Protected areas have been proposed by some
authors as the only solution to the overfishing problems (Birkeland, 1997;
Russ, 1966) and reserves may be the only way to provide critical refuges for
large and long-lived species (Huntsman, 1994) and those that undergo sex-change
during growth (like the groupers).
Sexually mature adult populations will be maintained in reserves and
larval export to other reefs will continue to provide a supply of juveniles
that can recruit to the fished population (see below).
Marine reserves are
equally important in maintaining the genetic biodiversity of wild populations
of large fish species. These wild
populations are important to aquaculture of tropical marine fishes, which in
its infancy, relies on wild-caught broodstock.
The most fecund broodstock are the big ones that are genetically
selected for fast growth, disease resistance, and large maximum size. These big individuals are fished
preferentially for broodstock and are increasingly only available in reserves
or in lightly fished areas. In any
area, where fishing pressure is heavy enough to remove fish shortly after first
spawning, there is actually an active reduction in natural selection for the
attributes wanted by aquaculture broodstock.
High levels of exploitation of wild population do not allow the genetic
advantages of large final size to be expressed. This fisheries selection pressure towards earlier reproduction
and smaller maximum size has been demonstrated for populations in the Caribbean
and the Grand Banks (Man et al.., 1995).
Marine reserves are
important for protecting the spawning stock of natural populations and
broodstock for aquaculture. In
protecting these populations, the biological attributes of high fecundity and
fast-growth, stored in the genotype of big individuals, will be prevented from
being lost. High fecundity is
proportional to body size; the bigger the fish the greater is the
fecundity. However, the fecundity of a
big fish is not proportional to the weight of small individuals; a big fish
produces almost twice as many eggs as the same weight of small fish. For example, one 12.5-kg large snapper
produces the equivalent number of eggs as two hundred thirty three 1-kg
snappers (Bohnsack, 1994). Thus, it is
an evolutionary advantage that allows fish to grow as fast as possible to the
maximum size, which the environment can support.
Similarly, fast
growth is important in natural populations and in aquaculture. It limits the time an individual spends at a
size that is vulnerable to predation, allowing it to reach a large maximum size
and increasing fecundity levels. Having
large parents gives the maximum genetic advantage to offspring such as valuable
combinations of genes are passed on for rapid growth, large maximum size,
disease resistance, and other survival characteristics that enabled the parents
to attain large sizes in the hostile environment of the natural reef. Until there is a comprehensive series of
reserves that are linked by larval transport, there is a strong argument for
the adoption of a maximum size limit for exploitation as a way to protect some
of the genetic diversity of the largest fish species.
The other large
marine organisms that are heavily overfished and locally extinct are the eight
giant clam (Tridacnidae) species which have been overfished for meat and shells
throughout their tropical Indo-Pacific distribution (Lukas, 1994). The last 30 years have been particularly
severe for the two largest species, due in part to international poaching for
their adductor muscle. Low densities,
erratic recruitment, and a relatively slow growth rate to reach harvestable
size make these populations prone to overfishing. Even on the remote reefs of Layang Layang, population sizes for T.
maxima indicate that the population is recovering from heavy fishing
pressure in the past. Evidence of fishing
pressure on stocks can be seen on most Indo-Pacific islands from the mean size
of clams near a village compared with distant parts of the lagoon (Braley,
1989). In many areas, populations of T.
squamosa are critically low and T. maxima are locally extinct. Declining giant clam stocks and local
extinction's were a major stimulus for research into mariculture methods and
there are several commercial farms now in operation.
Marine parks are
important in conserving the genetic variability within populations if the
population as a whole is to survive.
Fished populations are especially at risk from genetic selection
pressure; for example, size-selection of gear is a strong selective force for
fishes with a small size-at-maturity.
Furthermore, genetic variability is directly related to population size
and population size at maximum sustainable yield (MSY) is generally only half
the size of the unfished population.
Populations of many fish and invertebrate species on many reefs in the
South China Sea are well below MSY (McManus et al.., 1994) and are
therefore at risk from loss of genetic biodiversity. Conservation of populations is thus necessary but it is unknown
what minimum population size can maintain adequate genetic diversity and
minimise the risks associated with natural disasters or parasites. It is likely though that the minimum
population size is likely to vary between species to preserve genetic
variability.
Protection of biomass
It has now been
well established that marine reserves commonly support higher densities and
larger sizes of heavily fished species than are found outside reserves. The abundances and average sizes of many
larger carnivorous fishes are numerous and bigger within protected areas (Polunin
and Roberts, 1993). In areas where
smaller fishes are also targeted by fishers, species from all trophic levels
show similar responses to reduction in fishing pressure. In the 2 years following the set up of the
Saba Marine Park in the Netherlands Antilles, there was a 60% increase in overall
biomass of commercially important families while the predatory snappers
(Lutjanidae) increased by 220%. In
comparative studies of coral-reef fish communities of Saba Marine Park and Hol
Chan Marine Reserve (Ambergris Caye, Belize) in the Caribbean, 45 % of target
species commonly recorded in visual censuses in Belize, and 59% at Saba, showed
greater abundance, size or biomass in shallow protected sites (Polunin and
Roberts, 1993). The greatest estimated
biomasses were observed in locally protected snapper (Lutjanidae) in Belize and
Saba, and grunt (Haemulidae) at Saba.
In both protected areas, the local stock of visible demersal target
fishes was 1.9 to 2.0 times greater in biomass and 2.2 to 3.5 times greater in
commercial value than in fished sites.
Similarly, in reserves in the central Philippines, fish abundance's and
standing stock of highly fished species were significantly higher than in
non-reserves and control sites.
In other places, there is evidence that marine
reserves that allow limited fishing using traditional methods can still be
beneficial. On the Kenyan coast,
species richness was highest in marine parks where no fishing or collecting was
allowed but the same was not true for fish abundance, or for biomass of
commercially important fish (Samoilys, 1988).
Some of the highest densities and weights of fish were recorded from the
marine reserves where limited fishing using only traditional fishing methods
was allowed. Areas with higher fishing
intensity had smaller standing crops of fish, but not comparably smaller abundances.
Reserves clearly
create a local increase in abundance and size of fishes after the elimination
of fishing mortality, which in turn leads to greater egg production per unit of
reef and greater export via pelagic dispersal to fishing grounds as well as
spillover emigration to local reefs.
"Spillover" of individuals across reserve borders has been
shown for protected areas in the Philippines where there was a significantly
increased fish yield from nearby traditional fishing areas (White, 1989). There are good reasons to expect such
spillover from most reserves, and there is limited direct evidence for it
(Rowley, 1994). However, the magnitude
of any resulting increase in local catches will be difficult to predict and
very difficult to measure.
The actual species
that spillover will vary with behaviour.
Many small coral reef fish are strictly territorial and have small
ranges (Barrett, 1995). Territorial
ranges approximately 100 by 25-m or less were found in six wrasse, and
monocanthid species. The natural
habitat boundary of open sand between the reef and adjacent reefs appeared to
be an effective deterrent to emigration for these species. Other species notably the nocturnal squirrel
and soldier fish as well as the carangids, grunts, and snappers are swim over a
wide range and cross sandy areas of lagoon on a regular basis.
In general, field
studies from widespread sites around the globe support predictions of increases
in abundance and average size of fishes in protected areas but the manner in
which reserves enhance larger-scale recruitment to fish stocks is difficult to
quantify in the field. A
meta-population computer model shows how marine reserves help to conserve reef
fish populations and benefit fisheries (Man et al.., 1995). Simulations of the model show that reserves
become highly beneficial as the local extinction rate caused by fishing becomes
large because they provide a source of recruitment into fished-out patches. The abundance of the exploitable population
and the sustainable yield is maximised when half of all patches (reserves +
exploited patches) are occupied by the stock.
In such circumstances, the introduction of reserves meets the needs both
of conservation and of sustainable exploitation of the fishery.
Reserves can also
allow coral reef recovery after ecosystem damage by cyanide or dynamite
fishing. Reefs in the Philippines which have been protected after habitat
destruction show evidence of recolonisation by corals and other sedentary
benthic species (White, 1989).
Export of biomass and increase fishery yields
Marine
reserves are effective in maintaining high abundance in the reserve and higher
yields of outside of the reserve (e. g., Alcala and Russ, 1990; Russ and
Alcala, 1996). The proposed mechanism
for the increase in yields is by emigration of juveniles and adults from the
reserve to the fished site. Russ et
al.. (1994) simulated this mechanism in a yield-per-recruit model for
Sumilon Island Fishery Sanctuary, central Philippines. Later, circumstantial evidence of the export
of biomass was shown from Apo Island, central Philippines from a long-term
visual census data set and from data on fishery yields (Russ and Alcala, 1996).
To
make marine reserves effective to enhance fishery yields, the location of
marine reserves within the South East Asia region needs to be situated using
two distinct types of reserves. There
is a need for some areas to be located in middle/edge arrangement of biodiversity
reserves, linked to biogeographical regions (Hockey and Branch, 1994). Such reserves would achieve conservation of
both representativeness (middle) and high diversity areas (edge).
The sizes of
biodiversity reserves should be as large as possible, to allow for the
maintenance of populations of the largest reef species, and be located in areas
where current flow is reasonable, to allow the export products of larvae are as
widely dispersed as possible. There is
also a need for a second tier of small reserves which cover approximately 20 %
of all coral reef areas (McManus, 1995) for the specific purpose of
improving and maintaining local yields of exploited species. Second-tier smaller reserves should be
designed to maximise their benefit to adjacent areas while minimising their
size. Important populations can be
maintained in small reserves whose primary function is to supply recruits and
spillover of adults to nearby fished reefs.
Reserves of this nature would be ideal for giant clams were recommended
by several authors be established in each Indo-Pacific lagoon to ensure
continued reproductive success of the two most endangered species (Lucas,
1994). The other function of the
secondary reserves is as stepping-stones for maintenance of genetic biodiversity. Populations of most marine organisms in
minor reserves will be too small to maintain adequate genetic diversity, but
having a series of reserves which are all interlinked by larval transport to
the larger biodiversity reserves will limit loss of genetic information and
will almost eliminate permanent local extinction of species.
There are several
functioning examples of second tier reserves; the small island sites of Apo,
Negros and Sumilon, Cebu, in the Philippines both have fringing coral reefs and
support local fishers (White, 1989). In
the province of Negros Oriental, blastfishing has been stopped, cyanide fishing
in non-existent, and about 19 preserves are in place and working (CENTRO, pers.
comm.). The sites have a community
based marine resource management system.
With management, the coral reefs are a significant resource, providing
economic benefits that accrue to local fishers. These benefits include: (1) increased fish yields from
traditional fishing areas; (2) increased fish diversity and abundance within
sanctuary areas; (3) slightly improved coral substrate cover resulting from the
use of less damaging fishing methods; and (4) increasing tourism.
Marine Parks as
Sources of larvae: Evidence from Layang
Layang Island
Empirical evidence for
the important role of marine parks as sources of larvae and recruits are
limited. We report here a preliminary study of the role of good reefs in
exporting larvae to downstream reefs. The study was conducted at Layang Layang,
in the Spratlys, one of the reefs being claimed by Malaysia. It is distant from
population centres and the only infrastructures on the Island are the Royal
Malaysian Navy base and a dive resort.
The presence of the Royal Malaysian Navy and the inaccessibility of the
atoll result to the reefs not being fished actively and are isolated
effectively from all other human influences such as pollution and habitat
destruction. Thus, it remains a pristine, undamaged coral atoll with some of
the best coral reefs in the world (Mackey et al.., 1997). It also has
large spawning populations of many commercially important reef fishes and
clams.
Larvae were collected in the waters
arriving and leaving Layang Layang during the summer of 1996 (Oakley, unpub.
data; Figure 1). Plankton nets with 508 micron-mesh and diameter of 50 cm were
towed at 4 stations in the north, in the centre, on the reef, and in the south
of the Island. Stations 1 and 4 are approximately 500 m North or South of the
plankton stream. Stations 2 and 3 are in the plankton stream. Nets were towed
obliquely at 2 knots from 10 m to the surface at 2 knots for 10 minutes. Horizontal
tows were also at the same speed and duration.
Larvae were preserved in the field and later counted in the laboratory.
The pattern of
larval distribution in the waters around Layang Layang clearly demonstrated the
export of larvae from the reefs (Figure 1). In the waters arriving on the
atoll, there were almost no planktonic larvae of reef-based populations in the
water. As the water passed through the reef, larvae of many reef-based species
were released in the water, increasing the numbers of larvae in the water, and
then carried away. Thus, the departing
plankton stream has more larvae and could be clearly identified (Figure 2)
although the direction of the stream changed in relation to wind and water
currents (Figure 1). The majority of plankton were carried to the North West by
the prevailing current although on calm days there was an additional wind
driving surface flow to the East (Figure 3).
It is possible that the larvae that travelled eastward eventually mixed
with the predominant current and were carried back across the reef. This may be one of the mechanisms, which
returns larvae to their natal reef.
However, these larval export studies are only just beginning and more
information will be needed before larval export models can be developed for
Layang Layang.
These results from
the study on from Layang Layang Reef are empirical evidence on the important
role of protected reefs for fisheries although the Reef is not an established
marine park. In effect, however, Layang Layang Reef, is a marine park due to
the absence of exploitation and minimal developmental activity. The export larvae on the out-going tide and
current from the reef provides evidence of the mechanism by which good reefs
serve as sources of larvae.
Marine
parks for fisheries management
Marine reserves
have the potential to play an important role in fisheries management especially
in regions where there is heavy overfishing and ecosystem destruction. Enforcement of regulations in the region has
proved to be almost impossible, cyanide fishing for the live fish trade
(Johannes and Riepen, 1995) and dynamite are both banned but continue to be
used on a regular basis. The fishers often know the effects their fishing is
having but cannot stop because of the need to provide food and the feeling that
the common resource has to be taken before it disappears. Community based management with its own
enforcement can eliminate the problem of the common resource and can function
as an important step to conserve biodiversity and local areas. Unfortunately, protection of vulnerable
species and maintenance of resources is only likely to be successful if
networks of reserves are established throughout the region to link larval
supply and settlement areas.
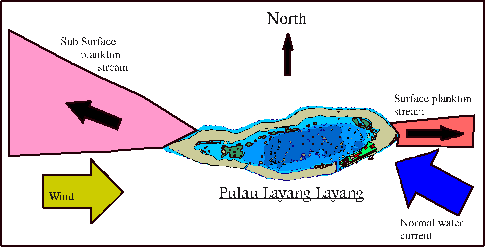
Figure 1.
Plankton exports during the summer months from Pulau Layang Layang in
the Malaysian Spratly Islands. The wind
driven surface flow to the E was only visible in calm sea conditions. Both plankton streams were clearly
detectable 1km from the reef.
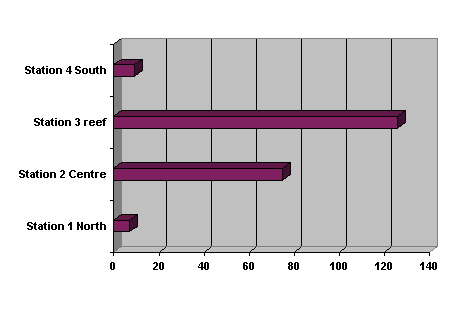
Figure 2.
Higher plankton concentrations (organisms per 100 ml) to the West of
Pulau Layang Layang in Stations 2 & 3 in the plankton stream and than in
Stations 1 & 4 that are approximately 500 m to N or South of it. Samples were from oblique tows in Stations
1, 2, and 4 are approximately 1 km W of reef while Station 3 is approximately
500 m W.
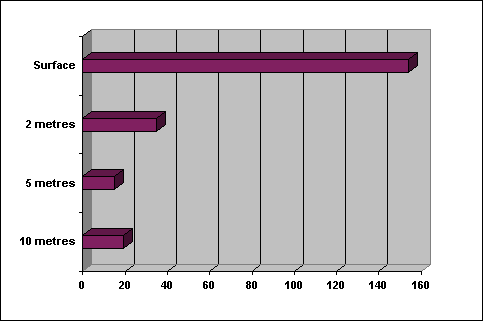
Figure 3. Decreasing
plankton concentrations (organisms per 100 ml) with depth to the East of Pulau
Layang Layang from horizontal plankton tows.
Samples were all taken in the middle of the stream.
Although
success with village-based marine reserves (White, 1989; Russ and Alcala, 1994)
in the Philippines indicates that local coastal management may be more
realistic than that based solely on national regulations, there remain some
problems in its implementation depending on the locality and situation. Problems
with marine parks in Indonesia and Papua New Guinea show that local systems
only work until there is foreign or industrial style fishing competing for the
same resource (Johannes and Riepen, 1995). Marine reserves are difficult to
establish and manage locally if the reefs in question are remote or
out-of-sight of the community (Arquiza and White, 1994).
Spratlys as a source for the reefs around the South China
Sea
The planktonic
larvae produced within the Spratly islands may be drifted around the South
China Sea (McManus, 1994). Using the speed and direction of predicted water
currents and from information on planktonic larval life, McManus (1994)
calculated that larvae from the Spratly islands could travel to many of the
heavily fished coastal reefs of the region.
This theory is based on the knowledge that planktonic or pelagic larvae
remain in the water column for periods that vary from a few days to a few weeks
(Thorson, 1971; Cameron, 1986) for most invertebrates although several
commercial lobster species have long planktonic lives (Sims and Ingle, 1966).
Coral reef fishes, for example, have a variable duration of planktonic and
pelagic phase but some do stay in the plankton for up to 100 days (Leis, 1991;
Brothers and Thresher, 1985). However, even for larvae of schooling species,
the eggs and early larval stages are almost certainly truly planktonic and a
significant proportion of eggs and larval fish are carried away from reefs.
The results of the study
from Layang Layang, in the Spratlys, provide empirical evidence or larval
export from the Spratlys. Furthermore,
it showed that the direction of the larval export could depend on the
prevailing monsoonal winds and current that prevail in the South China Sea. It
thus supports the hypothesis that the coral reefs in the Spratlys can be
sources of recruits for depleted reefs around this marginal sea (Figure 1).
Spratlys as a
Marine Park
The Spratlys, a
group of islands in the South China Sea, are surrounded by coral reefs that are
still in excellent condition (unpub. data). Brunei, China, Malaysia,
Philippines, Taiwan, and Vietnam are claiming these islands in whole or in
part. It is understood that China occupies currently seven of the Spratly
reefs, the Philippines around 12, the Vietnamese no less than 27 reefs,
Malaysia 4 reefs, and Taiwan and Brunei one each (Scholes, 1995). The belief
that the seabed adjacent to this group of islands and reefs contain oil and gas
and the strategic significance of the islands for sealane defence,
interdiction, and surveillance (Valencia et al.., 1994) drive the claims
to parts of the Spratlys.
Layang-Layang Reef, in the Spratly Islands, can serve
as the "source" of larvae and juveniles for reefs with depleted populations
of target species. The Reef has populations of common species at densities up
to 15 per 100 m2 that is higher than most reefs surveyed
by us (unpub. data). Large groupers are
still being caught in the vicinity of Layang Layang in the South China Sea. A
recent example was an individual, measuring over 3 m long and weighing
approximately 230 kg. This individual has the potential of carrying the genetic
potential for large size and the potential to produce eggs equivalent to 523
one-kg fish. A fish of this size are naturally rare in shallow depths (40 m and
shallower) and was not observed by us but by Filipinos who were known to fish
down to 60 m. Layang-Layang is thus worth conserving.
As a whole, the
Spratly Islands were considered worthy of conservation (Gomez, 1994; McManus,
1994). It is an ideal biodiversity reserve as it is in the centre of global
tropical marine biodiversity. It is ideally situated because no indigenous
population live in the region and it is fished except to satisfy the live fish
trade. A Marine Park for the Spratly Islands was already proposed earlier for
conservation (Gomez, 1994; McManus, 1994).
This arrangement requires a moratorium among the claimants in the
Spratlys and limits further development while various options to resolve the
conflict are being considered in the region. One of the proposed options is a
United Nations-sponsored marine protected area with zones and rights assigned
in a similar manner to the Great Barrier Reef Marine Park or Antarctica (Gomez,
1994; McManus, 1994).
With a marine park
in the Spratlys, regulatory measures on fishing may be implemented to conserve
the fishery resources that threatened by overfishing and habitat-destruction. Environmentally
destructive fishing methods, currently uses, need to be banned and less
destructive methods can be used to harvest the productivity of the Spratly
reefs. Indeed, if we could persuade
fishers to target medium-sized fishes, we could harvest a far greater biomass
(10 to 20 times) for the same effect on the fecundity of the population
(Birkeland, pers. comm.). Fishers could catch to the maximum sustainable yield
without affecting population genetic potential since most species are
recruitment-limited. This is a strong argument for not catching a few large
individuals that could produce as many eggs as possible and enhance
recruitment. Fortunately, protective management for coral reefs has this and
other several advantages. Hopefully it will not be essential to fully
understand the variable nature of recruitment and yield relationships before
steps can be taken towards establishing a series of protected areas where fish
stocks can recover from overfishing and habitat destruction.
Benefits of Marine Park in the Spratlys to Fisheries around the
South China Sea
Larval export from
reserves or unfished reefs like Layang Layang in the Spratlys has the potential
to supply recruits to reef fisheries over large regions, like the South China
Sea. The success of marine parks for fisheries enhancement will depend upon
many factors, including the knowledge of movement patterns and habitat
requirements of all life stages (larval, settlement, juvenile, adult, feeding,
and breeding) for each targeted species.
This is clearly a difficult task when there are over 300 species of fish
and invertebrates represented in Southeast Asian fish markets. As a step
towards this goal, information on relative rates of production inside and
outside reserves (Bohnsack, 1993), and patterns of larval transport and
mortality, are being collected by the UNIMAS project to determine whether they
may play a role in enhancing or sustaining catches in fished areas. Egg production in reserves and non-reserve
areas will be estimated from knowledge of population structures and behaviour
of the species concerned. The information from these studies will be useful in
designing more effective marine reserves and to demonstrate conclusively the
value of reserves on fisheries.
Coral reef areas
with high standing stock of targeted species are important in replenishing not
reefs open to fishing but also those reefs protected from fishing. Even reefs
that are protected from fishing, such as the Saba Marine Park (Netherlands
Antilles), has low population densities of groupers, perhaps due to a lack of
supply of larvae from unprotected source areas (Roberts, 1995). Clearly, a
"source" reef, like Layang Layang, is important in replenishing
viable populations even in protected areas.
More marine parks needed
Marine parks are important and beneficial not only for
tourism and education. These parks play an important role in aquaculture and
fisheries. In order to retard the impacts of anthropogenic uses of the reefs,
particularly exploitation of living resources, more marine parks need to be
established to for sustainable fisheries.
Acknowledgement
We are deeply indebted to the
Royal Malaysian Navy, for permission and logistical support to conduct research
on Pulau Layang Layang. Sarawak Shell
Sdn. Bhd., our long- term supporters of marine conservation at UNIMAS, has
contributed financially towards this research on Pulau Layang Layang. Others who have contributed include the
Layang Layang Resort, the team members from the University of Aberdeen 1996
Expedition and Esso Malaysia Sdn. Bhd.
This work could not have been completed without the support and
encouragement of Prof. Ghazally Ismail, Deputy Vice Chancellor Universiti
Malaysia Sarawak. This work was partly
funded by UNIMAS research grant 89/96(8), for which we are very grateful.
Literature
Cited
Alcala, A. C. 1981.
Fish yields of coral reef of Sumilon Island, Central Philippines. Nat. Res. Counc. Philipp. Bull. 36:1-7.
Alcala,
A. C. 1988. Effects of marine reserves on coral reef fish abundances and
yields of Philippine coral reefs. Ambio
17:194-199.
Alcala, A. C. and G. R. Russ. 1990.
A direct test of the effects of protective management on abundance and
yield of tropical marine resources. J.
Cons. Int. Explor. Mer. 46:40-47.
Alcala, A. C. and F. J. Vande Vusse. 1994.
The role of government in coastal resources management. pp. 12-19.
In: Pomeroy, R. S.
(ed.) Community management and common
property of coastal fisheries in Asia and the Pacific: concepts, methods, and experiences. ICLARM Conf. Proc. 45. 189 pp.
Arquiza,
Y. and A. White. 1994. Tales from Tubbataha. Bandillo ng Palawan Foundation, Inc., Puerto
Princesa, Palawan. 136 pp.
Barrett,
N.S. 1995 Short and long term movement patterns of six temperate reef fishes
(Families Labridae and Monacanthidae).
Aust. J. Mar. Freshw. Res.
46(5): 853-860.
Birkeland, C. E.
1997. Life and death of coral
reefs (ed.). London: Chapman &
Hall, 527 pp.
Bohnsack,
J. A. 1993. Marine reserves.They enhance fisheries, reduce conflicts, and
protect resources. Oceanus 36(3): 63-71.
Bohnsack,
J. A. 1994. How marine fishery reserves can improve fisheries. Pp 217-241.
In Proc. 43rd Conf.
Gulf and Caribbean Fisheries Institute, Miami, Florida, 1990.
Braley,
R. D. 1989. A giant clam stock survey and preliminary investigation of pearl
oyster resources in the Tokelau Islands.
ACIAR Giant Clam Proj., James Cook University of North Queensland,
Townsville, Australia, 90 pp.
Brothers,
E. B. and R. E. Thresher. 1985. Pelagic duration, dispersal, and the
distribution of Indo-Pacific coral reef fishes. In: Reaka, M.L. (ed.).
The Ecology of Coral Reefs.
Symposia Series for Undersea Research, NOAA's Undersea Research Program,
3(1). p. 53-69.
Cameron,
R. A. 1986. Reproduction, larval occurrence, and recruitment in Caribbean sea
urchins. Bull. Mar.
Sci. 39:332-346.
Cordner,
L. G. 1994. The Spratly Islands dispute and the Law of the Sea. Ocean Dev.
Int’l. Law 25(1): 61-74.
Chou,
L. M. 1997. The status of Southeast Asian coral reefs. Pp. 317-322. In: Lessios, H. A
and I. G. Macintyre. Proc. 8th
Intl. Coral Reef. Symp. Panama: Smithsonian Tropical Research Institute.
Dixon,
J. A. 1993. Economic benefits of marine protected areas. Oceanus 36(3):35-40.
Dugan,
J. E. and G. E. Davis. 1993 (eds).
Applications of marine refugia to coastal fisheries management. Int. Symp. Marine Harvest Refugia, San
Antonio, Texas, USA, 12 Sep 1991.
Dugan,
J. E. and G. E. Davis. Applications of
marine refugia to coastal fisherires management. Can. J. Fish. Aquat. Sci. 50( 9): 2029-2042.
Gomez,
E. D. 1994. The South China Sea – conservation are or war zone ? Mar. Poll. Bull. 28(3):132.
Hockey,
P. A. R. and G. M. Branch. 1994.
Conserving marine biodiversity on the African coast: Implications of a
terrestrial perspective. Aquat. Conserv.
Mar. Freshw. Ecosys. 4(4): 345-362.
Huntsman,
G. R. 1994. Endangered marine finfish: Neglected resources or beasts of
fiction? Fisheries 19(7): 8-15.
Johannes,
R. E. and M. Riepen. 1995. Environmental, economic and social
implications of the live reef fish trade in Asia and the West Pacific. The Nature Conservancy, Honolulu, Hawaii. 87
pp.
Leis,
J. M. 1991. The pelagic stage of reef fishes: the larval biology of coral
reef fishes. pp. 183-230.
In: The Ecology of Fishes on
Coral Reefs. Sale, P. F. (ed.). San Diego, California: Academic Press,
Inc. 754 pp.
Lucas,
J. S. 1994. The biology, exploitation, and mariculture of giant clams
(Tridacnidae). Rev. Fish. Sci. 2(3): 181-224.
Mackey,
G., R . Clubb, C. Digges, S.
Enderby, M. Keller, and K.
Atack. 1997. Final Report on the Joint University of
Aberdeen, University of Malaysia Sarawak, Expedition to Pulau Layang Layang
1996.
Man,
A., R. Law, and N. V. C. Polunin. 1995.
Role of marine reserves in recruitment to reef fisheries: A
metapopulation model. Biol. Conserv.
71(2):197-204.
McDorman,
T. L. 1993. The South China Sea islands dispute in the 1990s a new
multilateral process and continuing friction.
Int. J. Mar.
Coast. Law 8(2):263-285.
McManus,
J. W. 1994 . The Spratly Islands: A marine park. Ambio 23(3): 181-186.
McManus,
J. W. 1995a. Future prospects for artificial reefs in the Philippines. pp. 33-29. In: Munro, J.L. and P. E. Munro
(eds). Workshop on Artificial Reefs in
the Philippines, Manila (Philippines), 30-31 Aug 1994. Manila, Philippines. ICLARM Conf. Proc.
49. pp. 33-39.
McManus, J. W. 1995b.
Large and small scale marine protected areas: Planning, investment and
intergenerational quality of life. pp.
60-71. In: Hooten, A. J. and M. E. Hatziolos (eds). Sustainable Financing Mechanisms for Coral
Reef Conservation Workshop, Washington, DC, USA, 23 June 1995. Environ. Sust. Dev. Proc. Ser. 9.
McManus,
J. W. 1997. 1997. Tropical marine
fisheries and the future of coral reefs: a brief review with emphasis on
Southeast Asia. Pp. 129-134. In:
Lessios, H. A and I. G. Macintyre.
Proc. 8th Intl. Coral Reef. Symp. Panama: Smithsonian
Tropical Research Institute.
McManus,
J. W., C. L. Nañola, Jr., R. B. Reyes, Jr., and K. N. Kesner. 1994.
Resource Ecology of the Bolinao Coral Reef System. ICLARM Studies and Reviews 22, 117 p.
Oakley,
S. G. 1984. The effects of spearfishing pressure on grouper (Serranidae) populations in the Eastern
Red Sea. Pp. 341-359 In: Proc. Symp.Coral Reef Env. Red Sea. Faculty of Marine Science, King Abdul Aziz
University, Jeddah, Saudi Arabia.
Polunin,
N. V. C. and C. M. Roberts. 1993. Greater biomass and value of target coral
reef fishes in two small Caribbean marine reserves. Mar. Ecol. Prog. Ser. 100:167 - 176.
Roberts,
C. M. 1995. Rapid build up of fish biomass in a Caribbean marine
reserves. Conserv. Biol. 9(4): 816-826.
Roberts,
C. M. and N. V. C. Polunin.
1993. Marine reserves: Simple
solutions to managing complex fisheries? Ambio 22(6): 363-368.
Rowley,
R. J. 1994. Marine reserves in fisheries management. Aquat. Conserv. Mar. Freshwat. Ecosyst. 4(3):233-254.
Rubec, P. J.
1988. The need for conservation
and management of Philippine coral reefs. Envir. Biol. Fishes. 23: 141-154.
Russ,
G. R. 1996. Fisheries management: what chance on coral reefs? NAGA 19(3):5-9.
Russ,
G. R. and A. C. Alcala. 1989. Effects of intense fishing pressure on an
assemblage of coral reef fishes. Mar.
Ecol. Prog. Ser. 56:13-27.
Russ,
G. R. and A. C. Alcala. 1994.
Sumilon Island reserve: 20 years of hopes and frustrations. NAGA 17(3) 8-12.
Russ,
G. R. and A. C. Alcala. 1996. Do marine reserves export adult fish biomass
? Evidence from Apo Island, central
Philippines. Mar. Ecol. Prog. Ser.
132:1-9.
Russ, G. R., A. C. Alcala, and A. S.
Cabanban. 1994. Marine reserves and
fisheries management on coral reefs with preliminary modelling of the effects
on yield-per-recruit. In: Proceed. 7th Intl. Coral Reef Symp.
1:988-995.
Sadovy,
Y. 1996. Reproduction of reef fishery species. pp. 15-60. In
N.V.C. Polunin and C.M. Roberts (eds).
Reef Fisheries. London: Chapman and Hall. 477 pp.
Samoilys, M. A.
1988. Abundance and species
richness of coral reef fish on the Kenyan coast: The effects of protective
management and fishing. pp.
261-266. In: Choat, J. H., D.
Barnes, M. A. Borowitzka, J. C. Coll, P. J. Davies, P. Flood, B. G. Hatcher, D.
Hopley, et al.. (eds). Vol. 2.
Proceedings of the Sixth International Coral Reef Symposium, Townsville,
Australia, 8th -12th August 1988.
Scholes,
W. 1995. Escalating tension over disputed Spratly Islands. Pet.
Rev. 49(581):262.
Sims,
H. W. and R. M. Ingle. 1966.
Caribbean recruitment of Florida’s spiny lobster population. Quarterly Jour. Flor. Acad. Sci. 29(3):206-242.
Thorson,
G. 1971. Life in the Sea. New
York: McGraw Hill Book Company. 256 pp.
Trippel,
E. A. 1995. Age at maturity as a stress
indicator in fisheries. BioScience 45
(11):759-771.
Valencia,
M. J. and, D. Zha. 1994.
The South China Sea disputes: Recent developments. Ocean. Coast.
Manage. 24(3):199-204.
White, A. T.
1989. Two community based marine
reserves: Lessons for coastal management. pp. 85-96. In: Chua, T. E. and D. Pauly (eds). Coastal Area Management in Southeast Asia: Policies, Management
Strategies and Case Studies. ICLARM
Conf. Proc. 19